Comment le QI peut être déterminé par la génétique ? La génétique passe d’abord par l’ADN et l’ARN, contenant l’information génétique, cela se passe dans le noyau, une fois un gène transcrit ce sont les protéines qui ont un effet sur l’organisme. Cette protéine peut agir à plusieurs niveaux. Au niveau de l’intelligence, différents systèmes biologiques sont concernés. L’intelligence est corrélée avec :
- La vitesse de traitement de l’information,
- La taille du cerveau,
- L’épaisseur de la matière grise,
- L’intégrité de la matière blanche entre deux régions corticales.
- La composition neuronale.
Le cerveau est donc composé de neurones. Le nombre de neurones et l’intelligence ont une corrélation positive. Ces neurones communiquent au sein du cerveau, ainsi vient la synapse, la rapidité du message synaptique a une corrélation positive avec l’intelligence. La synapse participe également à la plasticité cérébrale, à l’adaptation du cerveau au changement. Le cerveau est aussi un organe qui est le siège de dynamiques ; la projection et différentiation neuronale et des oligodendrocytes qui participe à la myélinisation du système nerveux central qui permet une conduction rapide du message nerveux le long des synapses. Ainsi, à chaque étape, des gènes interviennent pour activer, inhiber, des réparations peuvent être faites, des rétrocontrôles positifs ou négatifs ont lieu lors de ces événements, c’est un bouillonnement de matière. Le nombre et le type de protéines influençant le développement et la dynamique cérébrale sont déterminés par l’ADN.
Etudes sur le score polygénique.
Les premières études de Davide Piffer sur la génomique ont permis de débroussailler le terrain. Les avancées en biologie moléculaire permettent de faire avancer la science en général, le domaine du QI n’y échappe pas. (Piffer 2014; 2015)
La sélection faiblement répandue ou l’adaptation polygénique est un mécanisme agissant sur plusieurs SNP. C’est un modèle proposé pour expliquer l’évolution de traits hautement polygéniques, déterminés par une variation génétique commune et ancienne. (Pritchard et Di Rienzo 2010)
L’étude propose une démarche dans la méthode de détection des signaux de sélection polygénique sur des gènes dont les phénotypes se chevauchent, malgré leurs différences de positions chromosomiques. La méthode est donc différente des estimations traditionnelles du déséquilibre de liaison que nous verrons par la suite. Le but de l’étude est de suggérer une méthodologie pour détecter les signaux de sélection polygénique en utilisant le niveau de scolarité. C’est ce qu’il nous intéresse.
Le niveau de scolarité est un phénotype polygénique influencé par de nombreuses variantes génétiques ayant de faibles effets individuels. Les fréquences alléliques sont positivement corrélées aux mesures globales du niveau de scolarité et du QI moyen. Les allèles augmentant le QI seront positivement corrélés aux allèles augmentant le niveau de scolarité. Deux SNP dont les associations avec l’intelligence semblent robustes, car ils ont été répliqués dans plusieurs études indépendantes. Le premier est rs236330, situé dans le gène FNBP1L, dont l’association significative avec l’intelligence générale a été rapportée dans deux études distinctes.(Davies et al. 2011)
(Benyamin et al. 2014)
Ce gène est fortement exprimé dans les neurones, notamment les neurones de l’hippocampe et des cerveaux en développement, il régule la morphologie neuronale. Le deuxième est le rs324650. Il a été inclus, car son association avec le QI a été reproduite dans quatre études d’association. (Comings et al. 2003) (Dick et al. 2007) (F. M. Gosso et al. 2007; M. F. Gosso et al. 2006)
Il est localisé dans le gène CHRM2, récepteur muscarinique, impliqué dans l’excitabilité neuronale, la plasticité synaptique et la régulation par rétroaction de la libération d’acétylcholine.
Les pays du monde diffèrent dans leurs niveaux de réussite scolaire appréciable soit par la durée, soit par les diplômes, soit par les résultats scolaires. Plusieurs facteurs ont été avancés pour expliquer ces différences. Le plus souvent, cela a été attribué à des facteurs économiques et socioculturels, cependant, il n’est pas évident que la croissance économique conduit à des résultats plus élevés aux tests de réussite scolaire. (Hanushek et Woessmann 2010)
De plus, ce qui est souvent avancé néglige les variations génétiques entre les individus et les groupes humains, bien que la plupart des traits humains soient héréditaires dans une large mesure et que le niveau de scolarité ne fait pas exception. (Plomin et al 2008)
L’intelligence est un bon prédicteur de performance aux tests de réussite scolaire, en particulier dans des matières telles que les mathématiques où elle explique 58,6% de la variance dans une étude longitudinale portant sur plus de 70.000 enfants anglais, les experts ont trouvé des corrélations élevées entre les mesures du g académique et du g cognitif. (Deary et al. 2007) (Kaufman et al. 2012).
Le classement national des scores d’intelligence reflète étroitement le gradient observé dans les fréquences des gènes associés à cette construction. Les asiatiques de l’Est ont les fréquences les plus élevées d’allèles bénéfiques pour le niveau de scolarité (39%) et surpassent systématiquement tous les autres groupes ethniques. Les européens ont des fréquences légèrement inférieures (35,5%) et les Africains ont les fréquences d’allèles associées le plus faible (16%).
Les scores factoriels de 8 groupes raciaux étaient fortement corrélés à leur QI estimé (r=0.95). Il est important de noter que cette analyse réfute également toute affirmation selon laquelle les scores factoriels représentent simplement la distance de l’Afrique. Des pressions sélectives en faveur d’un QI plus élevé se sont poursuivies après la scission entre les populations d’Asie du Nord-Est et les amérindiens ou entre les populations d’Asie du Sud Est et les Océaniens.
Les populations d’Asie centrale et du Moyen-Orient ont des scores factoriels comparables à ceux européens, ce qui suggère que leur QI moyen plus faible pourrait être amélioré grâce à des conditions qui amélioreraient leur condition génétique et leur développement.
Les allèles associés à l’intelligence sont uniques aux humains et non partagés avec les primates non-humains. Ce résultat est prédit sur la base de l’hypothèse selon laquelle les humains ont évolué par sélection naturelle pour devenir plus intelligents que leurs cousins primates. Les résultats montrent que ce processus évolutif s’est poursuivi dans les populations humaines modernes. Cela invalide donc les théories qui supposent que l’évolution cognitive d’Homo sapiens a pris fin avec la première apparition de l’Homo sapiens.
La méthode d’analyse factorielle peut être utilisée comme outil de découverte de gènes. Les SNP hautement corrélés au score factoriel peuvent être utilisés dans de futures études d’association à l’échelle du génome. Cette stratégie peut réduire considérablement le nombre de SNP inclus dans une étude d’association à l’échelle du génome, réduisant ainsi le problème des tests multiples et la taille des échantillons requis. Ici, il s’agissait surtout à fournir des preuves systématiques que les différences entre les pays et les ethniques en matière de QI et de niveau d’éducation sont liées à des facteurs génétiques.
GWAS hits and intelligence
Si le QI a bien une détermination génétique, comment cela se produit ? Comment cela est construit ? Par exemple, les ashkénazes qui représentent moins de 10 millions de personnes à travers le monde représentent pourtant 50% des champions du monde d’échecs et de bridge et au moins 25% des prix Nobels mondiaux. Les ashkénazes ont un Q.I moyen encore supérieur à 110. L’argument environnemental ne tient pas, les juifs sont éparpillés dans le monde entier dans des systèmes bio-culturels différents. (Dunkel et al. 2019)
Dans l’ère du progrès de la biologie moléculaire, le séquençage complet du génome humain a été achevé en 2004, il y a un intérêt particulier chez les chercheurs d’isoler des gènes, à partir du génome humain, plutôt de l’exome, pour tenter d’expliquer leurs effets. Ainsi, pour les maladies l’intérêt est de coder l’exome du malade et de le comparer à un témoin contrôle pour comparer l’absence, la présence, ou la modification des gènes. Ainsi, les études GWAS (Genome Wide Association Study) sur l’intelligence (2015, 2018, 2019) ont été récemment réalisée par plusieurs chercheurs. Ces études se penchent sur l’ensemble des variations génétiques augmentant l’intelligence découverte à ce jour permettant la mise en lumière de nombreux variants génétiques différents impliqués dans un même trait phénotypique.
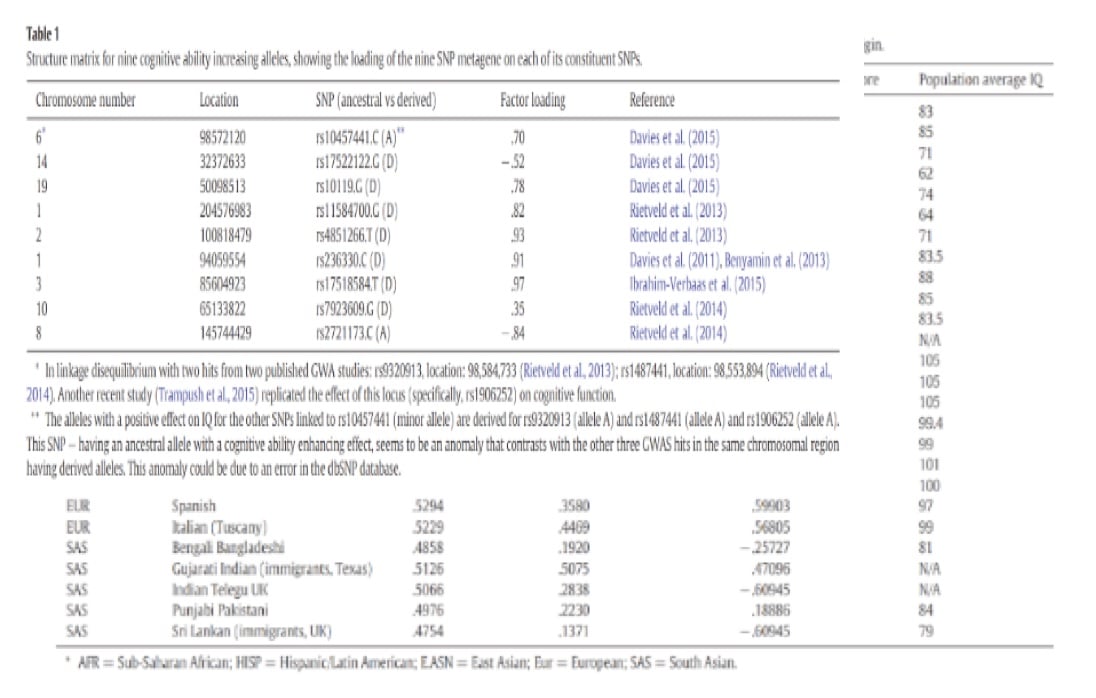
Ces études ont permis d’identifier une association d’allèles fortement en lien avec l’intelligence et le niveau d’éducation. La méta-analyse de Rietvelda identifié 10 polymorphismes mononucléotidiques qui ont augmenté le niveau de scolarité, comprenant trois ayant une signification nominale à l’échelle du génome et sept ayant une signification suggestive. (Rietveld et al. 2013)
Une récente étude (Ward et al. 2014) a reproduit l’effet positif de ces trois principaux SNP (rs9320913, rs11584700 et rs4851266) sur les mathématiques ainsi que sur les performances en lecture dans un échantillon indépendant d’écoliers. Ces SNP se sont également révélés associés à g (rappel : intelligence générale) dans un sous-échantillon de l’étude originale de Rietveld.
Ce tableau 1 est un peu compliqué à expliquer dans le détail. Il s’agit de la liste des SNP (polymorphisme nucléotidique) avec la location sur le chromosome. Il y a le facteur de corrélation qui évalue la puissance de corrélation entre le SNP et l’intelligence.
Le score polygénique est la moyenne des fréquences alléliques et les QI moyens de la population sont présentés dans le tableau 2. Il y a une corrélation positive et significative entre les neuf SNP métagène et QI (r = 0,863, N = 23). C’est la méthode des vecteurs de corrélation qui a été utilisée pour évaluer l’impact prédictif de la validité de l’analyse factorielle.
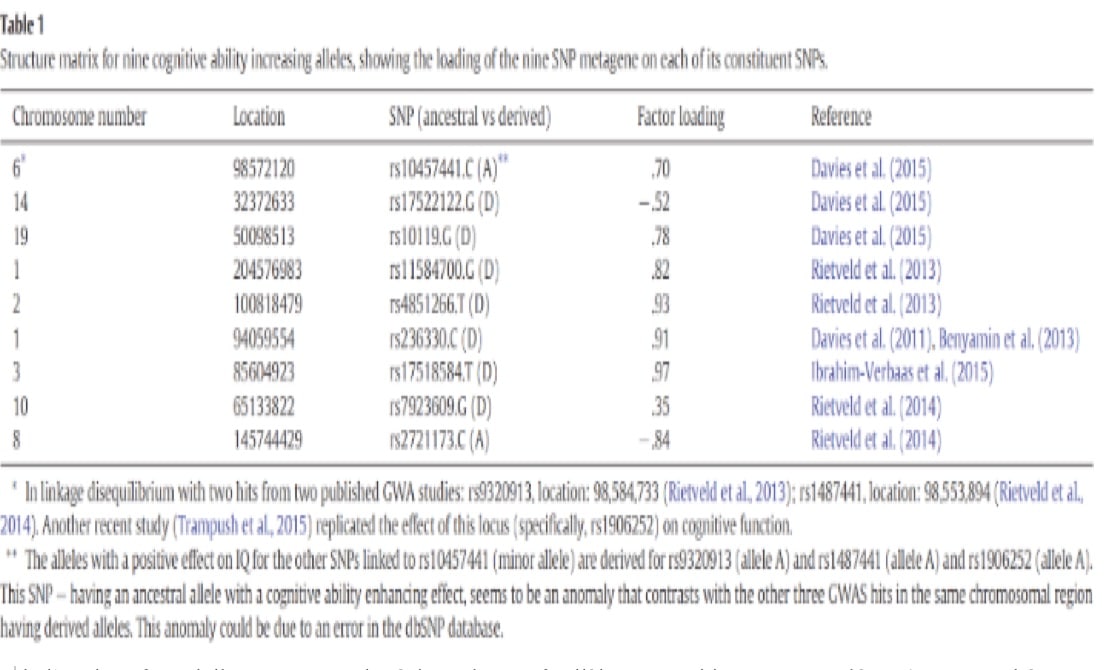
Le score polygénique des neufs allèles est positivement associé au QI et aux phénotypes. La comparaison des moyennes de fréquence des allèles pour les cinq groupes continentaux de la base de données 1000 Genomes a révélé des différences de fréquence qui correspondent étroitement au QI global observé au niveau du continent, donnant le modèle suivant : Asie de l’Est > Europe > Asiatique du Sud > Américain (hispanique) > Africain.
Dans ce tableau ci-dessous, nous voyons bien que le QI national est corrélé positivement au score polygénique. Nous remarquons que pour certaines ethnies, un score polygénique ne suit pas parfaitement le QI. Il y a toujours une corrélation positive, mais la valeur de corrélation varie en fonction de la population concernée. Nous expliquerons cette variation par la suite.
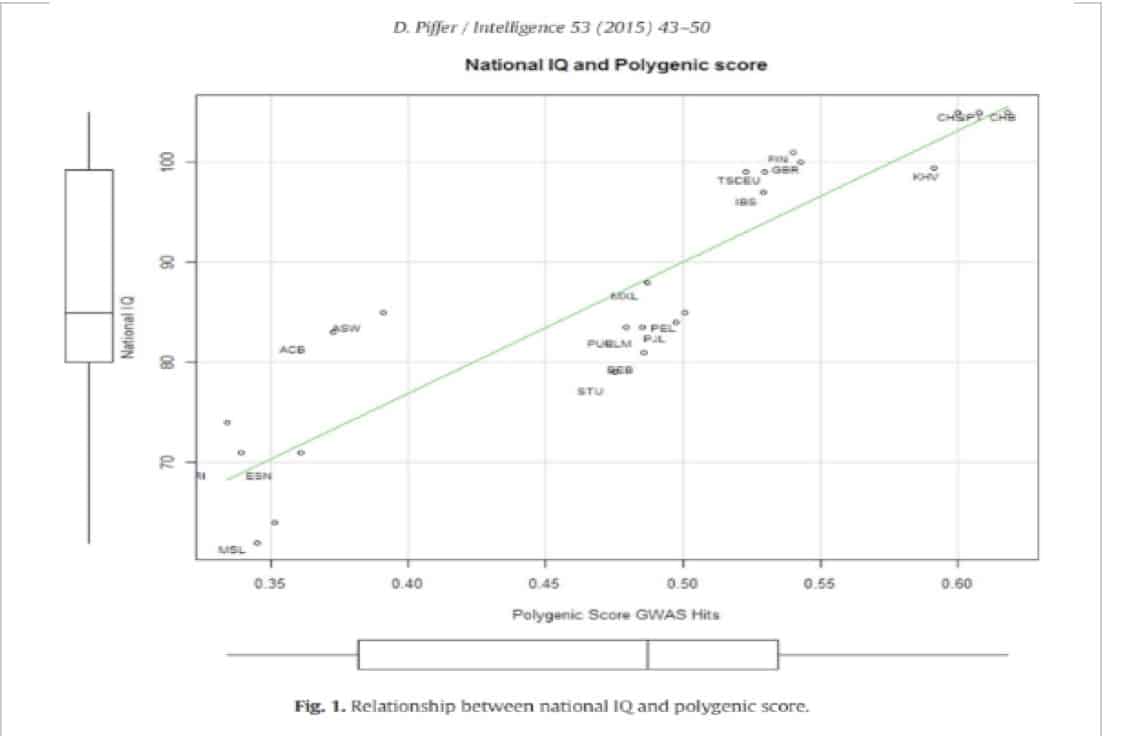
Cependant, il convient de dire, sans que cela contredise tous les points de ce passage, que l’ANOVA n’a pas donné p valeurs qui répondent au seuil de signification conventionnel de .05. De plus le test de Tukey a produit des intervalles de confiance qui ont divisé zéro en deux. Le manque de signification statistique est clairement dû au très petit échantillon taille (N = 9). Il est donc nécessaire de continuer les recherches sur ce sujet, il fait sans nul doute que le nombre croissant d’études GWAS fourniront davantage de résultats à l’avenir, permettant ainsi de générer une étude de plus en plus approfondie et une image de plus en plus précise de la variation génétique liée à la cognition, à la fois au sein et entre les populations.
En comparant les fréquences moyennes de la taille et d’intelligence, une différence frappante est la variabilité inter-population beaucoup plus élevée de cette dernière, allant de 33,4% (Luhya, Kenyan) à 61,82% (Han Chinese, Pékin). À l’inverse, les fréquences moyennes des allèles augmentant la taille varient de 44,76 % (vietnamiens) à 49,5 % (Esan nigérian). (Piffer 2015)
Ainsi, on a seulement 5% de différences de score polygénique alors que pour l’intelligence, la différence est de près 30%. Cela semble indiquer que le signal de sélection est beaucoup plus fort pour l’intelligence que la taille, créant ainsi une grande variabilité entre les populations. Une piste pouvant expliquer cette différence est que les facteurs environnementaux ou sociaux (culturels, sexuels…) qui affectent les différences de conditions liées à l’intelligence varient davantage que les conditions liées à la taille. Cet écart pourrait être boosté par la disparité des conditions lié à l’intelligence. La détermination des différences est génétique mais l’environnement pourrait moduler cette différence.
D’ailleurs, la distinction entre sélection directionnelle et diversification est pertinente à cet égard. Ainsi, une intelligence « génotypique » plus élevée pourrait provoquer le développement de société plus complexe et la vie dans ces sociétés complexes sélectionnerait des gènes à QI élevé. Clark avait proposé des mécanismes de coévolution culture-gène pour expliquer l’augmentation du capital humain parmi les populations préindustrielles d’Angleterre. Nous pouvons aussi supposer que l’effet Flynn a permis aux populations européennes de vivres dans des sociétés plus complexes dans leur organisation. (Clark et Hamilton 2006)
Les recherches récentes sur l’évolution de la taille et de l’intelligence ont apporté des résultats encourageants. En effet, il présente des preuves d’une dominance directionnelle sur la taille et l’intelligence (mais pas sur d’autres traits comme la tension artérielle ou le cholestérol des lipoprotéines de basse densité). Cette étude fournit donc la preuve qu’une stature et une capacité cognitive accrues ont été positivement sélectionnées dans l’évolution humaine.
(Joshi et al. 2015)
Cette sélection se résume à une association de neufs allèles. Les neuf allèles sont toutes présentes à des niveaux significatifs, fréquences (>5%) parmi les cinq grandes races (Afrique subsaharienne, Asiatiques du Sud, Européens, Asiatiques de l’Est, Américains) (voir Tableau 8). Ainsi, les polymorphismes intellectuels ne semblent pas spécifiques à la race comme elles étaient déjà présents chez Homo sapiens avant l’exode africain vers 60-100 Kya. De plus, il ne semble avoir un polymorphisme spécifique à l’Europe (Tableau 8). Il est donc probable que la grande majorité des mutations affectant l’intelligence étaient déjà présentes dans la population ancestrale africaine et à mesure que les humains se sont installés dans différentes parties du monde, ces polymorphismes étaient soumis à une pression de sélection directionnelle, qui a produit une augmentation globale de l’intelligence humaine à des rythmes différents selon les zones géographiques. Notons qu’il peut y avoir des effets pléiotropes sur des traits autres que le QI, de sorte que les gènes du QI soient soumis à d’autres types de pression de sélection. L’analyse d’un plus grand nombre de gènes semble nécessaire, car les effets confondants des gènes pléiotropes s’annuleront au fil du temps et un grand nombre d’allèles s’exprimeront dans différentes orientations pour différents phénotypes par la pression de sélection.
Ainsi, il faudrait que les recherches continuent, avec notamment la combinaison avec les statistiques de Wright, dont F(ST), qui fournissent des informations sur les processus évolutifs qui influencent la structure des variations génétiques entre les populations. Ces estimations peuvent identifier des régions du génome qui ont été la cible de la sélection, et les comparaisons de F(ST) provenant de différentes parties du génome peuvent fournir des informations sur l’histoire démographique. Il est souvent estimé à partir des polymorphismes mononucléotidiques, qui peut caractériser une sélection directionnelle. Ses valeurs varient de 0 (panmixie) à 1 (différenciation complète). (Holsinger et Weir 2009)
Par exemple, les valeurs de F (ST) dépendent fortement des populations. Il y a des groupes ethniques étroitement liés, tels que le Danois et les Néerlandais, ou le Portugais et l’Espagnol afficher des valeurs de FST nettement inférieures à 0,01 (1%), indiscernables de la panmixie. En Europe, les groupes ethniques les plus divergents se sont révélés avoir des valeurs de l’ordre de 0,07 (7%) (Lapps et Sardes).
Des valeurs plus importantes sont trouvées si des groupes très purs génétiquement sont trouvés. C’est le cas pour certaines peuplades d’Afriques et d’Océanie qui ont vécus plus ou moins en autarcie, généralement des tribus n’acceptant pas le métissage avec d’autres. Ainsi, la valeur la plus élevée est 0,46 (46%) entre les Mbuti et Papous. (Cavalli-Sforza, Menozzi, et Piazza 1995) (Ginsburgh et Weber 2016)
Petite application souvent difficile à se le représenter, la parenté entre le parent et la progéniture de 1/4 décrit le partage de gènes au-delà du partage aléatoire dans une population d’accouplement aléatoire. Dans une population subdivisée, la statistique Fst décrit le partage des gènes au sein des subdivisions de la même manière. Une distance génétique de 0,125 ; c’est-à-dire la moyenne de 10 et 15%, qui est l’intervalle moyen de proportion FST, implique que la parenté entre des individus non apparentés de la même ascendance par rapport à la population mondiale équivaut à une parenté entre demi-frères/sœurs, ou grands-parents/petit-enfant, dans une population d’accouplement aléatoire. Cela implique également que si un humain d’une population ancestrale donnée à un demi-frère mixte, cet humain est génétiquement plus proche d’un individu indépendant de sa population ancestrale que de son demi-frère mixte. (Harpending 2002)
La génétique moléculaire des populations est un domaine en pleine expansion et dans un avenir proche, davantage de gènes liés aux capacités cognitives seront, sans aucun doute, identifié.
Alternativement, sur une échelle de temps plus petite, les récentes tendances dysgéniques en matière d’intelligence dans les sociétés industrielles peuvent être quantifiées au niveau moléculaire, corroborant ainsi d’éventuelles preuves phénotypiques (Woodley of Menie 2015) (Menie et al. 2015)
Ces études ont évalué ensuite les différences ethniques dans la fréquence de ces allèles, et ont montré qu’elles étaient strictement parallèles aux différences de Q.I entre populations ; En d’autres termes, les ethnies à Q.I plus élevé ont effectivement une fréquence supérieure d’allèles augmentant l’intelligence dans leur patrimoine génétique.
GWAS-hits : une réplication des découvertes précédentes.
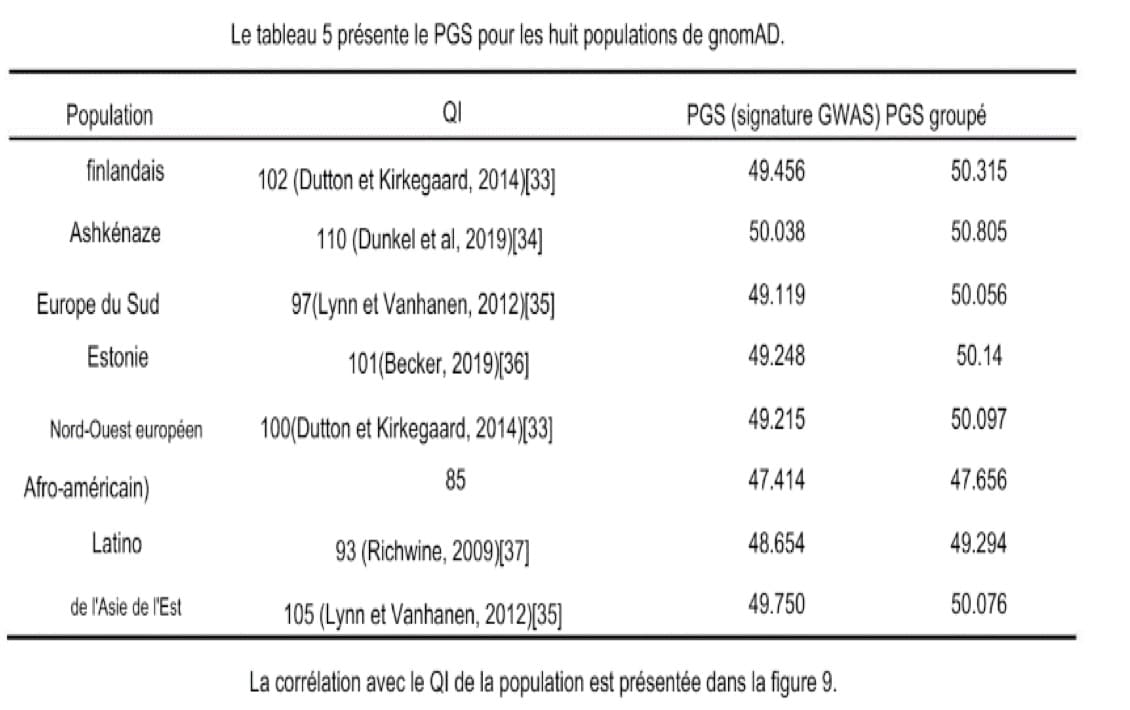
L’étude menée par D. Piffer publié en 2019 permet de confirmer les conclusions des découvertes précédentes. (Piffer 2019). Il a déjà été abordé plus haut qu’il pourrait avoir eu une récente sélection sur l’intelligence. Les GWAS-hits permettent de rendre compte de ce phénomène, et c’est dans cette étude que nous pouvons voir les progrès sur ce sujet. Ainsi, dans cette étude il y a l’utilisation des variants génétiques identifiées par les trois études d’association pangénomique (GWAS) pour tester un modèle de sélection polygénique liée à l’intelligence. L’étude prend le départ sur les scores polygéniques (PGS) qui sont comparés entre les populations à l’aide des données ; 1000Genomes, HGDP-CEPH et gnomAD. Au sein des échantillons gnomAD, la corrélation entre PGS et QI est presque parfaite (r=0.89, N=8) et l’ANOVA a montré des différences significatives entre les populations dans les fréquences alléliques avec un effet positif. L’environnement, les variables socio-économiques ont légèrement amélioré la précision du modèle (environ de 80% à 85%), mais le PGS a expliqué deux fois plus de variances du QI par rapport aux variables socio-économiques.
Les généticiens des populations ont reconnu que la plupart des caractères sont hautement polygéniques et ont abandonné l’étude de l’évolution génétique sous l’aspect mendélien monogénique pour utiliser des modèles examinant plusieurs gènes ensemble. Les recherches sur les variations de caractères complexes ont montré des différences significatives. L’identification de ces différences et des adaptations polygéniques est compliquée par plusieurs facteurs.
La présence de milliers de variants génétiques, chacun ayant un faible effet, impliquant que les signaux de différenciation génétique sont dilués et non-évidents et qu’il faut des méthodes de balayages de plus en plus sensibles.
L’identification d’un nombre de SNP suffisant pour expliquer au moins 5% de la variance totale d’un caractère nécessite de grands échantillons et ceux-ci ne sont devenus disponible que récemment.
Les facteurs environnementaux, différant selon les populations, peuvent influencer le phénotype et masque partiellement les différenciations génétiques.
L’un des objectifs de cette étude est de reproduire les effets découverts par Piffer, en utilisant les preuves du nouveau GWAS sur le lien entre l’intelligence et le niveau de scolarité publié à ce jour, lien par des facteurs hautement prédictif du QI de la population. La méthode d’analyse factorielle repose sur l’hypothèse que la sélection polygénique agit comme une variable latente qui explique les points communs entre plusieurs variants génétiques dispersées au sein du génome. Ce modèle intègre un terme d’erreur ; erreurs de mesure, de couverture imparfaite ou de dérivé génétique, qui augmentent le bruit.
La limite principale des scores polygéniques est que les études GWAS sont réalisées majoritairement sur des populations européennes. Cela pose quelques problèmes :
- Le GWAS ne parviendra pas à capturer les variantes spécifiques à une population.
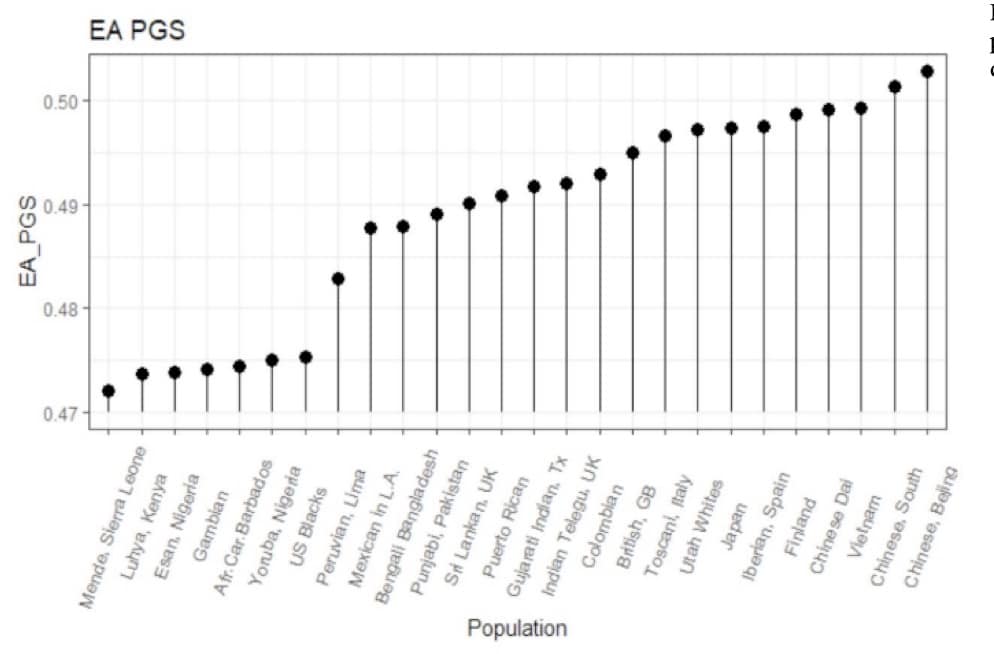
Cela ne biaise nécessairement le PGS en faveur du groupe de référence, car le GWAS identifie les variants d’effets négatifs et positifs. Par exemple, un récent GWAS réalisé sur des Péruviens a trouvé une variante spécifique qui réduit la taille d’environ 2,2 cm. Etant donné que cette variante n’est polymorphe que dans les populations amérindiennes, elle aurait été manquante dans un GWAS sur des populations européennes.
Le calcul des scores polygéniques au niveau de la population est une approche prometteuse et rapide pour tester les signaux d’adaptation polygénique. Les résultats démontrent clairement des différences de populations en termes de PGS en corrélation avec les estimations du QI moyen de la population, et les performances des élèves aux tests standardisés de mathématiques, de lecture et de sciences (r= .9 et .8)
La grande réplicabilité du même modèle polygénique issu d’une publication précédente (Piffer 2015) confère davantage de force aux présents résultats. Piffer a calculé les scores polygéniques et factoriels pour une population de 1000 génomes en utilisant les données disponibles à l’époque de GWAS avec une taille d’échantillon beaucoup plus petit. Ainsi, cette étude renforce grandement les estimations précédentes des PGS très élevés (r= 0.95 à 0.98) en corrélations avec les estimations du QI moyen.
Les scores plus élevés de l’Asie de l’Est suggèrent qu’une pression de sélection s’est poursuivie après la scission entre l’Asie de l’Est et les Amérindiens, environ 15.000 plus tôt. La date de la migration à travers le détroit de Béring vers l’Asie est contestée. Il y a des preuves de la paléogénomiques qui démontre que la scission s’est produite il y a 20.000 ans alors que les deux résidaient encore en Asie du Nord-Est, et qu’il y a eu un flux génétique continu jusqu’à ce que le groupe amérindien traverse le détroit de Béring vers l’Amérique du Nord vers 15.000 ans. Il est possible que la densité de population extrêmement faible dans les Amériques ait réduit la compétition intraspécifique, diminuant donc la sélection sur les capacités cognitives était donc plus faible. Sujet ouvert au débat.
Une des limites de la présente étude réside dans le fait qu’elle s’appuie sur des estimations du QI de la population comme variable, phénotypique et parfaitement imprécises ; ce sont des moyennes et ils peuvent, en partie, refléter les différences environnementales et économiques entre les populations. De plus, l’EA GWAS pourrait capturer la variation génétique qui contribue au niveau de scolarité via des mécanismes autres que le QI. Il est aussi connu que les scores polygéniques fonctionnement mieux dans les populations européennes et que la précision des prédictions est réduite d’environ 2 à 5 fois dans les populations d’Asie de l’Est et d’Afro-américaine, respectivement. (Martin et al. 2019)
En 2018, une étude de Lee a constaté sur un échantillon d’Afro-américains qu’effectivement EA PGS plus élevé était associé à une probabilité plus élevée d’achèvement des études universitaires et de performances en mathématiques, mais pas à la réussite en lecture. (Lee et al. 2018)
Qu’il était aussi associé à une association négative avec le casier judiciaire. Cependant, il y a eu une atténuation par rapport à l’effet PGS chez les populations de races blanches. Les auteurs attribuent cela à divers facteurs potentiels :
- L’étude a utilisé un échantillon de faible statut socio-économique, où l’environnement partagé joue un rôle plus important que dans les environnements à SES élevé. (Menie et al. 2015)
- Les mesures de performances en mathématiques et en lecture ont été obtenus dans la petite enfance, période de développement au cours de laquelle l’héritabilité, l’influence génétiques sur l’intelligence est moindre (et celle de l’environnement plus élevée) par rapport aux jeunes adultes.
Cela montre bien que les scores polygéniques sont plus précis pour les peuples blancs, il y a un euro-centrisme. Cependant, bien que moins précis, ces résultats restent pertinents. Il faut simplement continuer les études et séquencer la génétique des autres races.
De plus, les méta-analyses transethniques GWAS sur d’autres traits ont également trouvé des variantes génétiques avec peu d’hétérogénéité entre les groupes d’ascendance. (Lee et al. 2018)
Un récent GWAS sur la schizophrénie a révélé qu’environ 95% des SNP d’un Occidental, blanc évidemment, avaient une direction d’effet cohérente dans l’échantillon chinois. (Pietschnig et Voracek 2015)
Ainsi, on comprend donc que les imprécisions sur les autres races en raison de l’euro-centrisme n’annulent pas évidemment les études relatives à cela. En effet, les êtres humains étant tout de même assez proches génétiquement, le score éloigné de FST n’est « que » de 46%, même s’il y a cet euro-centrisme, ce manque de précision n’implique pas des différences drastiquement différentes.
Ainsi, ce noyau commun de variantes génétiques causales ayant des effets trans-ethniquement homogènes est ce qui détermine l’association entre les valeurs moyennes des traits et le QI ou la taille de la population, ainsi que les différences de groupe dans le PGS moyen.
La fréquence de l’allèle SNP moyen est de 50%, les SNP auront tendance à converger vers une fréquence moyenne. Cette fréquence de 50% avec une décroissance croissante du LD. L’implication de cela pour notre analyse est que lorsque les scores polygéniques sont inférieurs à 50%, nos estimations seront gonflées, et idem pour les PGS > 50%. En effet, la décroissance des LD fait monter (ou diminuer) les scores polygéniques vers la fréquence de fond de 0.5. Dans le cas présent, la fréquence moyenne des allèles à effet positif est de l’ordre de 50% (49,7%et 50,1%) pour les CEU et CHS.
CEU et CHS font référence à deux populations spécifiques qui ont été largement utilisées dans des études de génétique des populations et de génomique. CEU : « Utah residents with Northern and Western European ancestry ». CHS : « Han Chinese South ».
Ici, la décroissance du LD ne devrait produire qu’un léger biais dans l’estimation. Cependant, les fréquences alléliques des SNP causals dans d’autres populations (non blanche) plus éloignées de 50% produiront un biais plus fort. Par exemple, la moyenne de l’YRI (population noire) est de 47,5% donc la décroissance du LD produit une surestimation du PGS. La fréquence des SNP causals devrait être inférieure à 47.5%.
L’étude de Lee démontre bien que plus la fréquence est faible, plus l’estimation est élevée dans l’ensemble plus large que dans l’ensemble des SNP présumés causals, ceci suggère bien que la décroissance des LD conduit à une surestimation du PGS dans les populations avec un PGS plus faible, comme prévu par la théorie. (Lee et al. 2018)
L’ajout de variables socio-économiques au modèle a légèrement augmenté le pouvoir prédictif (de 80% à 85%), bien que le PGS explique deux fois plus la variance (70% contre 35%) que ceux de l’IDH, de la consommation moyenne de protéines ou de la mortalité infantile. L’inverse était le cas pour la taille où les facteurs SES expliquaient beaucoup plus la variance de la taille moyenne que le PGS. Ceci est conforme aux études d’héritabilité qui montrent l’importance de l’environnement partagé pour la taille adulte, même dans les pays occidentaux riches (environ 10%) et plus encore dans les pays non-occidentaux. (Flynn 2011) D’un autre côté l’impact environnemental partagé sur le QI à l’âge adulte s’avère généralement proche de 0. (Lynn et Irwing 2004).
Bien que cela ne soit pas forcément le cas dans les pays en développement où l’environnement joue un rôle plus important que les pays occidentaux. En effet, le QI des Afro-américains semble être plus élevé que ce qui est prédit par le PGS, ce qui suggère que cela ne peut pas être expliqué par le seul mélange européen, mais pourrait être le résultat d’une meilleure nutrition ou d’infrastructures éducatives par rapport aux autochtones africains. Une autre explication est l’hétérosis, la vigueur hybride, c’est-à-dire l’augmentation de la condition physique observée chez la progéniture hybride grâce à l’expression réduite d’allèles récessifs délétères homozygotes. (Shull 1948)
Pour les recherches futures, une voie intéressante serait de tester des modèles plus sophistiqués avec des ensembles plus larges de variables socio-économiques. Les futures études GWAS devraient être réalisées sur des populations non-européennes, car les GAWS transethniques constituent une ressource prometteuse pour l’identification d’allèles ayant des effets homogènes et hétérogènes ainsi que pour le calcul de PGS spécifiques à une population. Cela nous permettrait d’inclure les SNP polymorphes uniquement dans certaines populations et de trouver les SNP causals communs à l’homme.
Zoom sur le national QI.
Dans un preprint, Rebecca Sear en mai 2022 apporte une nuance sur les valeurs de « National QI » rapportés par Lynn et Vanhanen. (Sear 2022) Les données rapportées par Lynn et Vanhanen en 2002 ont été beaucoup utilisé et selon Rebecca Sear leur utilisation est impropre. L’étude évalue la dernière version, produite en 2019, par Lynn et Becker, et tend à démontrer que les données sont inadaptées à son objectif. (Lynn et Becker 2019) Rebecca Sear critique des échantillons non représentatifs, une méthode de tests cognitifs différents et auprès de populations si diverses qu’il est impossible de générer une mesure comparable de la capacité cognitive et qu’ainsi toutes les études utilisant ces données ne doivent pas être pris en compte et mis de côté.
Les estimations des scores du QI national ont engendré une controverse depuis leur première publication en 2002. Une grande partie des chercheurs ont utilisé ces scores moyens pour identifier les relations entre le national QI et les variables économiques et sanitaires, une autre partie ont soutenu que les valeurs du score du QI national sont sans fondement et que toute étude qui les utilise est irrémédiablement erronée. L’étude de Russel Warne a pour but d’évaluer la qualité des estimations du national QI, de discuter de la validité des différentes interprétations et suggérer des solutions susceptibles de compenser les lacunes des données sous-tendant la moyenne estimée. (R. Warne 2022) Tout cela pour permettre à la communauté scientifique de rejeter la dichotomie et l’absence de réserve. Que l’on peut voir dans l’article de Rebecca Sear. Au cours des décennies, les estimations du national QI ont été mises à jour à plusieurs reprises. (Lynn et Meisenberg 2010) (Lynn et Vanhanen 2006) (Lynn et Vanhanen 2012). La dernière version date de 2019. (« View on IQ » 2019) En conséquence, bon nombre des critiques formulées à l’encontre des versions antérieures de la base de données en raison de l’inclusion d’échantillons spécifiques est moins pertinent aujourd’hui. En effet, bon nombre des échantillons les plus critiqués ne sont plus inclus dans la version la plus récente. Les années de travail et l’accumulation des données ont permis de recueillir plus d’informations afin d’avoir des résultats plus proches de la réalité. Ces résultats sont aujourd’hui plus précis que ceux d’avant, bien qu’il reste inexact en absolue, ils permettent de mieux apprécier les différences de QI et de permettre une base de travail qu’il convient d’affiner.
L’étude dont il est question ici a pour but de générer des prédictions découlant de l’étude de Warne (R. T. Warne 2021) qui plaide en faveur de causes en partie génétiques des différences d’intelligence générale (g) entre les Américains auto-identifiés par la race ou l’origine ethnique (SIRE). Les tests de QI sont des prédicteurs statistiquement robustes de la réussite scolaire et professionnelle, et les différences de scores cognitifs moyens entre les groupes raciales/ethniques aux Etats-Unis ont été bien documentés. (Murray 2021) (Roth et al. 2017). Ces différences sont indues à un biais psychométrique, comme en témoigne l’invariance des mesures dans les groupes américains SIRE. (Frisby et Beaujean 2015) (Scheiber 2016). Les différences de scores représentent les différences moyennes réelles dans la construction de l’intelligence générale. Ainsi, l’écrasante majorité des experts attribue une partie des différences à la génétique. Les désaccords proviennent dans le degré de la contribution génétique. (Rindermann, Becker, et Coyle 2020) Le défi actuel est donc de déterminer la source de ces différences afin de mieux comprendre et d’aborder ainsi leurs implications sociales. (Flynn 2018) (Pesta, Fuerst, et Shibaev 2021).
Ainsi, Warne a décrit des preuves convergentes en faveur d’un modèle partiellement génétique pour les différences raciales :
- La conclusion cohérente de l’invariance.
- La découverte de l’héritabilité élevée au sein d’un groupe suggère que cela en est de même pour les différences entre groupes.
- Les résultats d’études basées sur la génétique utilisant les scores polygéniques
- Les résultats d’études basées sur la génétique appliquant « regression methodology ».
- Support cohérent à grande échelle basé sur la méthode des vecteurs corrélés.
Les experts de l’article suivent l’appel à la recherche de Warne. Pour cela, il y a d’abord l’étude de Warne. (R. T. Warne 2021) et ceux des auteurs cités par Warne : (Lasker 2019 ; Rushton et Jensen 2005 ; R. T. Warne 2020a ; 2020b)
A partir de cette analyse, il y a la construction de plusieurs arguments testés sur une grande base de données. La base de données exclut les personnes d’ascendances principalement asiatique ou insulaire du pacifique pour permettre d’avoir suffisamment de puissance statistique par la taille des échantillons concentrés surtout sur les ascendances européennes, Amérindiennes et Africaines. Les ensembles de données utilisés ont été 1000Genomes et HapMap.
L’hypothèse 1 : Il y a des différences moyenne, de modéré à élever, entre les groupes SIRE américains en intelligence, eduPGS et volume cérébral/intracrânien. Les données ont montré que c’était fortement le cas. De plus, notre analyse MGCFA a confirmé l’hypothèse de Spearman, selon laquelle les différences cognitives sont principalement dues à des différences de g. Cette découverte augmente l’hypothèse d’une forte incidence de la génétique, car les différences g sont plus héréditaires au sein des groupes SIRE.
L’hypothèse 3 : Les héritabilités sont similaires entre les groupes. C’est ce que l’on retrouve dans cette étude ; avec l’échantillon national d’enfants de dix ans les estimations de l’héritabilité de l’intelligence générale étaient modérées à élevées dans l’échantillon blanc et non-blancs. De plus, cela est conforme aux conclusions de Pesta. (Pesta et al. 2019 ; 2023) qui rapportent que les héritabilités sont similaires entre les groupes blancs et non-blancs.
L’hypothèse 5 : Les différences dans l’intelligence générale sont dues à l’ascendance génétique. Cela s’est avéré être le cas, conformément aux résultats de recherches récentes. (Fuerst, Kirkegaard, et Piffer 2021 ; Kirkegaard et al. 2019 ; Lasker et al. 2019 ; R. T. Warne 2020a ; 2020b) Dans l’échantillon complet, les variables n’avaient pas de validité indépendamment de l’ascendance génétique. De plus, l’ascendance européenne était positivement liée à g, tant que l’ascendance amérindienne et africaine l’était négativement. Cela implique que les grands facteurs socioculturels (ségrégation, discrimination, normes culturelles) ne sont pas à l’origine des lacunes en matière d’intelligence entre les groupes.
L’hypothèse 6 : Le score eduPGS prédit l’intelligence au sein des groupes SIRE et au sein des fratries. Cela s’est avéré être le cas. Conformément aux résultats précédents. (Howe et al. 2022) Nous avons constaté une contraction du PGS au sein de la fratrie, par rapport à l’association au sein de l’échantillon complet. Cette découverte conforte cette conclusion.
L’hypothèse 10 était de savoir si le SSE serait lié au g et au volume cérébral chez les enfants biologiques, mais pas dans les sous-échantillons d’enfants adoptifs. Il a été consté que le SSE n’était pas corrélé au g et au volume cérébral chez les enfants adoptés alors qu’il était positivement corrélé aux deux variables chez les enfants biologiques. Bien que ces résultats ne soient pas particulièrement solides, en raison de la petite taille et du manque d’informations préalables, ils sont cohérents avec ceux rapportés précédemment. Ceux qui montrent que le SSE des parents n’est que faiblement lié au QI de l’enfant adoptif. (McGue et al. 2007 ; Beaver et al. 2014). Les résultats actuels suggèrent provisoirement que la relation entre le SSE parental et g, dans cet échantillon, est principalement génétique.
National QI et PIB
De nombreuses publications ont mis en évidence une relation entre le QI et la prospérité nationale depuis la publication du livre « IQ and the Wealth of Nations ». (Lynn et Vanhanen 2002). L’étude d’E. Kirkegaard et G. Francis (Francis et Kirkegaard 2022) ont pour but d’étendre les travaux de Jones et Schneider (2006). (Jones et Schneider 2006). Cette étude constate que le QI national est le meilleur prédicteur de la croissance économique. Les meilleures estimations montrent qu’une augmentation d’un point du QI est associée à une augmentation de 7,8% du PIB par habitant.
Ces résultats ont des implications pour les politiques publiques et la croissance économique. Les faibles preuves venant confirmer la théorie « smart fraction » suggère que la présence d’une élite intellectuelle n’a que de faibles effets sur le PIB. Cela affaiblit les arguments en faveur des politiques qui tendent à mettre en place une élite en place pour concevoir et diriger des institutions clés dans les pays en développement. Ces résultats pourraient aussi indiquer que l’immigration peut réduire le PIB par habitant. Si un pays au QI élevé accueille des migrants au QI plus faible, la nouvelle moyenne peut déterminer la prospérité de la société, malgré la stabilité du QI de l’élite locale, et aussi baisser le QI moyen des deux nations par le départ des plus intelligents.
Les experts concluent que l’ensemble de la société devrait veiller à accroître l’intelligence. C’est une demande que les experts ont déjà exprimée, dont Cattell, un des psychologues les plus prolifiques, appela en 1937 les nations à développer des stratégies visant à accroître leur intelligence. (Cattell 1937) (« The Fight for Our National Intelligence / by Raymond B. Cattell. » s. d.)
Ainsi ; il faut avoir une discussion sur les implications éthiques et politiques du génie génétique. (Anomaly 2020).
De plus, comme le QI détermine le PIB, et donc les gènes et le score polygéniques, il faut s’attendre à une baisse de la croissance économique. Récemment, des données génétiques américaines. (Beauchamp 2016), du Royaume-Uni (Hugh-Jones et Abdellaoui 2021) et de l’Islande (Kong et al. 2017) montrent que les scores polygéniques (PGS) lié à l’intelligence sont en baisse.
Mazhev Roder-Heraod
Bibliographie et sources
Anomaly, Jonathan. 2020. Creating Future People: The Ethics of Genetic Enhancement. Taylor & Francis. https://doi.org/10.4324/9781003014805.
Beauchamp, Jonathan P. 2016. « Genetic evidence for natural selection in humans in the contemporary United States ». Proceedings of the National Academy of Sciences 113 (28): 7774 79. https://doi.org/10.1073/pnas.1600398113.
Beaver, Kevin, Joseph Schwartz, Mohammed Al-Ghamdi, Ahmed Kobeisy, Curtis Dunkel, et Dimitri Linden. 2014. « A closer look at the role of parenting-related influences on verbal intelligence over the life course: Results from an adoption-based research design ». Intelligence 46 (septembre): 179 87. https://doi.org/10.1016/j.intell.2014.06.002.
Benyamin, B., Bst Pourcain, O. S. Davis, G. Davies, N. K. Hansell, M.-J. A. Brion, R. M. Kirkpatrick, et al. 2014. « Childhood Intelligence Is Heritable, Highly Polygenic and Associated with FNBP1L ». Molecular Psychiatry 19 (2): 253 58. https://doi.org/10.1038/mp.2012.184.
Cattell, Raymond B. 1937. « PSYCHOLOGY AND SOCIAL PROGRESS: MANKIND AND DESTINY FROM THE STANDPOINT OF A SCIENTIST ». The Journal of Nervous and Mental Disease 86 (1): 108. https://journals.lww.com/jonmd/Citation/1937/07000/PSYCHOLOGY_AND_SOCIAL_PROGRESS__MANKIND_AND.51.aspx.
Cavalli-Sforza, Luigi, Paolo Menozzi, et Alberto Piazza. 1995. « The History and Geography of Human Gene ». In Princeton, New Jersey, USA. Vol. 54. https://doi.org/10.2307/2058750.
Clark, Gregory, et Gillian Hamilton. 2006. « Survival of the Richest: The Malthusian Mechanism in Pre-Industrial England ». The Journal of Economic History 66 (3): 707 36. https://doi.org/10.1017/S0022050706000301.
Comings, D. E., S. Wu, M. Rostamkhani, M. McGue, W. G. Lacono, L. S.-C. Cheng, et J. P. MacMurray. 2003. « Role of the Cholinergic Muscarinic 2 Receptor (CHRM2) Gene in Cognition ». Molecular Psychiatry 8 (1): 10 11. https://doi.org/10.1038/sj.mp.4001095.
Davies, G., A. Tenesa, A. Payton, J. Yang, S. E. Harris, D. Liewald, X. Ke, et al. 2011. « Genome-Wide Association Studies Establish That Human Intelligence Is Highly Heritable and Polygenic ». Molecular Psychiatry 16 (10): 996 1005. https://doi.org/10.1038/mp.2011.85.
Deary, Ian J., Steve Strand, Pauline Smith, et Cres Fernandes. 2007. « Intelligence and educational achievement ». Intelligence 35 (1): 13 21. https://doi.org/10.1016/j.intell.2006.02.001.
Dick, Danielle M., Fazil Aliev, John Kramer, Jen C. Wang, Anthony Hinrichs, Sarah Bertelsen, Sam Kuperman, et al. 2007. « Association of CHRM2 with IQ: Converging Evidence for a Gene Influencing Intelligence ». Behavior Genetics 37 (2): 265 72. https://doi.org/10.1007/s10519-006-9131-2.
Dunkel, Curtis S., Michael A. Woodley Of Menie, Jonatan Pallesen, et Emil O. W. Kirkegaard. 2019. « Polygenic Scores Mediate the Jewish Phenotypic Advantage in Educational Attainment and Cognitive Ability Compared with Catholics and Lutherans. » Evolutionary Behavioral Sciences 13 (4): 366 75. https://doi.org/10.1037/ebs0000158.
Flynn, James R. 2011. « Secular changes in intelligence ». In The Cambridge handbook of intelligence, 647 65. Cambridge handbooks in psychology. New York, NY, US: Cambridge University Press. https://doi.org/10.1017/CBO9780511977244.033.
———. 2018. « Academic freedom and race: You ought not to believe what you think may be true ». Journal of Criminal Justice, The Norman White Special Issue on the Study of Ethnicity and Race in Criminology and Criminal Justice, 59 (novembre): 127 31. https://doi.org/10.1016/j.jcrimjus.2017.05.010.
Francis, George, et Emil O. W. Kirkegaard. 2022. « National Intelligence and Economic Growth: A Bayesian Update ». The Mankind Quarterly 63 (septembre). https://doi.org/10.46469/mq.2022.63.1.2.
Frisby, Craig L., et A. Alexander Beaujean. 2015. « Testing Spearman’s hypotheses using a bi-factor model with WAIS-IV/WMS-IV standardization data ». Intelligence 51 (juillet): 79 97. https://doi.org/10.1016/j.intell.2015.04.007.
Fuerst, John, Emil O. W. Kirkegaard, et Davide Piffer. 2021. « More Research Needed: There is a Robust Causal vs. Confounding Problem for Intelligence-associated Polygenic Scores in Context to Admixed American Populations ». The Mankind Quarterly 62 (septembre): 151 85. https://doi.org/10.46469/mq.2021.62.1.10.
Ginsburgh, Victor, et Shlomo Weber, éd. 2016. The Palgrave Handbook of Economics and Language. London: Palgrave Macmillan UK. https://doi.org/10.1007/978-1-137-32505-1.
Gosso, Florencia M., Eco JC de Geus, Tinca JC Polderman, Dorret I. Boomsma, Danielle Posthuma, et Peter Heutink. 2007. « Exploring the functional role of the CHRM2 gene in human cognition: results from a dense genotyping and brain expression study ». BMC Medical Genetics 8 (1): 66. https://doi.org/10.1186/1471-2350-8-66.
Gosso, M. F., M. van Belzen, E. J. C. de Geus, J. C. Polderman, P. Heutink, D. I. Boomsma, et D. Posthuma. 2006. « Association between the CHRM2 Gene and Intelligence in a Sample of 304 Dutch Families ». Genes, Brain, and Behavior 5 (8): 577 84. https://doi.org/10.1111/j.1601-183X.2006.00211.x.
Hanushek, Eric A, et Ludger Woessmann. 2010. « The Economics of International Differences in Educational Achievement », mai.
Harpending, Henry. 2002. « Kinship and Population Subdivision ». Population and Environment 24 (novembre): 141 47. https://doi.org/10.1023/A:1020815420693.
Holsinger, Kent E., et Bruce S. Weir. 2009. « Genetics in Geographically Structured Populations: Defining, Estimating and Interpreting FST ». Nature Reviews Genetics 10 (9): 639 50. https://doi.org/10.1038/nrg2611.
Howe, Laurence J., Michel G. Nivard, Tim T. Morris, Ailin F. Hansen, Humaira Rasheed, Yoonsu Cho, Geetha Chittoor, et al. 2022. « Within-Sibship Genome-Wide Association Analyses Decrease Bias in Estimates of Direct Genetic Effects ». Nature Genetics 54 (5): 581 92. https://doi.org/10.1038/s41588-022-01062-7.
Hugh-Jones, David, et Abderrahman Abdellaoui. 2021. « Natural Selection in Contemporary Humans Is Linked to Income and Substitution Effects », janvier. https://researchinformation.amsterdamumc.org/en/publications/natural-selection-in-contemporary-humans-is-linked-to-income-and–2.
Jones, Garett, et W. Joel Schneider. 2006. « Intelligence, Human Capital, and Economic Growth: A Bayesian Averaging of Classical Estimates (BACE) Approach ». Journal of Economic Growth 11 (1): 71 93. https://doi.org/10.1007/s10887-006-7407-2.
Joshi, Peter K., Tonu Esko, Hannele Mattsson, Niina Eklund, Ilaria Gandin, Teresa Nutile, Anne U. Jackson, et al. 2015. « Directional Dominance on Stature and Cognition in Diverse Human Populations ». Nature 523 (7561): 459 62. https://doi.org/10.1038/nature14618.
Kaufman, Scott Barry, Matthew R. Reynolds, Xin Liu, Alan S. Kaufman, et Kevin S. McGrew. 2012. « Are cognitive g and academic achievement g one and the same g? An exploration on the Woodcock–Johnson and Kaufman tests ». Intelligence 40 (2): 123 38. https://doi.org/10.1016/j.intell.2012.01.009.
Kirkegaard, Emil O. W., Michael A. Woodley of Menie, Robert L. Williams, John Fuerst, et Gerhard Meisenberg. 2019. « Biogeographic Ancestry, Cognitive Ability and Socioeconomic Outcomes ». Psych 1 (1): 1 25. https://doi.org/10.3390/psych1010001.
Kong, Augustine, Michael L. Frigge, Gudmar Thorleifsson, Hreinn Stefansson, Alexander I. Young, Florian Zink, Gudrun A. Jonsdottir, et al. 2017. « Selection against variants in the genome associated with educational attainment ». Proceedings of the National Academy of Sciences 114 (5): E727 32. https://doi.org/10.1073/pnas.1612113114.
Lasker, Jordan. 2019. A Surfeit of Evidence on Spearman’s Hypothesis. https://doi.org/10.17605/OSF.IO/385AM.
Lasker, Jordan, Bryan J. Pesta, John G. R. Fuerst, et Emil O. W. Kirkegaard. 2019. « Global Ancestry and Cognitive Ability ». Psych 1 (1): 431 59. https://doi.org/10.3390/psych1010034.
Lee, James J., Robbee Wedow, Aysu Okbay, Edward Kong, Omeed Maghzian, Meghan Zacher, Tuan Anh Nguyen-Viet, et al. 2018. « Gene Discovery and Polygenic Prediction from a Genome-Wide Association Study of Educational Attainment in 1.1 Million Individuals ». Nature Genetics 50 (8): 1112 21. https://doi.org/10.1038/s41588-018-0147-3.
Lynn, Richard, et David Becker. 2019. The Intelligence of Nations.
Lynn, Richard, et Paul Irwing. 2004. « Sex differences on the progressive matrices: A meta-analysis ». Intelligence 32 (5): 481 98. https://doi.org/10.1016/j.intell.2004.06.008.
Lynn, Richard, et Gerhard Meisenberg. 2010. « National IQs calculated and validated for 108 nations ». Intelligence 38 (4): 353 60. https://doi.org/10.1016/j.intell.2010.04.007.
Lynn, Richard, et T Vanhanen. 2002. IQ and the Wealth of Nations.
Lynn, Richard, et Tatu Vanhanen. 2006. IQ and global inequality. IQ and global inequality. Augusta, GA, US: Washington Summit Publishers.
———. 2012. Intelligence: A unifying construct for the social sciences. Intelligence: A unifying construct for the social sciences. Ulster Institute for Social Research.
Martin, Alicia R., Masahiro Kanai, Yoichiro Kamatani, Yukinori Okada, Benjamin M. Neale, et Mark J. Daly. 2019. « Current clinical use of polygenic scores will risk exacerbating health disparities ». Nature genetics 51 (4): 584 91. https://doi.org/10.1038/s41588-019-0379-x.
McGue, Matt, Margaret Keyes, Anu Sharma, Irene Elkins, Lisa Legrand, Wendy Johnson, et William G. Iacono. 2007. « The Environments of Adopted and Non-Adopted Youth: Evidence on Range Restriction from the Sibling Interaction and Behavior Study (SIBS) ». Behavior Genetics 37 (3): 449 62. https://doi.org/10.1007/s10519-007-9142-7.
Menie, Michael A. Woodley of, Heitor B. F. Fernandes, Aurelio José Figueredo, et Gerhard Meisenberg. 2015. « By their words ye shall know them: Evidence of genetic selection against general intelligence and concurrent environmental enrichment in vocabulary usage since the mid 19th century ». Frontiers in Psychology 6. https://www.frontiersin.org/articles/10.3389/fpsyg.2015.00361.
Murray, Charles. 2021. « Facing Reality: Two Truths about Race in America – Murray, Charles: 9781641771979 – AbeBooks ». 2021. https://www.abebooks.com/9781641771979/Facing-Reality-Two-Truths-Race-1641771976/plp.
Pesta, Bryan, John Fuerst, et Vladimir Shibaev. 2021. Research Practices in Differential Psychology: Comments and a Correction to Winston. AZIMUTH OF SCIENTIFIC RESEARCH: PEDAGOGY AND PSYCHOLOGY. Vol. 10. https://doi.org/10.26140/anip-2021-1002-0098.
Pesta, Bryan, Emil O. W. Kirkegaard, Jan te Nijenhuis, Jordan Lasker, et John Fuerst. 2019. « Racial and ethnic group differences in the heritability of intelligence: A systematic review and meta-analysis ». Intelligence 78 (novembre). https://doi.org/10.1016/j.intell.2019.101408.
Pesta, Bryan, Jan te Nijenhuis, Jordan Lasker, Emil O. W. Kirkegaard, et John Fuerst. 2023. « On group differences in the heritability of intelligence: A reply to Giangrande and Turkheimer (2022) ». Intelligence 98 (mars): 101737. https://doi.org/10.1016/j.intell.2023.101737.
Pietschnig, Jakob, et Martin Voracek. 2015. « One Century of Global IQ Gains: A Formal Meta-Analysis of the Flynn Effect (1909–2013) ». Perspectives on Psychological Science 10 (3): 282 306. https://doi.org/10.1177/1745691615577701.
Piffer, Davide. 2014. « Factor Analysis of Population Allele Frequencies as a Simple, Novel Method of Detecting Signals of Recent Polygenic Selection: The Example of Educational Attainment and IQ ». The mankind quarterly 54 (janvier): 168 200. https://doi.org/10.46469/mq.2013.54.2.3.
———. 2015. « A review of intelligence GWAS hits: their relationship to country IQ and the issue of spatial autocorrelation. », janvier. https://doi.org/10.6084/M9.FIGSHARE.1461734.
———. 2019. « Evidence for Recent Polygenic Selection on Educational Attainment and Intelligence Inferred from Gwas Hits: A Replication of Previous Findings Using Recent Data ». Psych 1 (1): 55 75. https://doi.org/10.3390/psych1010005.
Plomin, R., et al. 2008. « [PDF] Behavioral Genetics (5th edition) Robert Plomin, John C. DeFries, Gerald E. McClearn, Peter McGuffin. (2008). Worth Publishers, New York, 505 pp., US$115.95, ISBN 10 1 4292 0577 6 | Semantic Scholar ». 1 avril 2008. https://www.semanticscholar.org/paper/Behavioral-Genetics-(5th-edition)-Robert-Plomin%2C-C.-Bates/f883eed97f37df5100f555413bcd552a64e28da0.
Pritchard, Jonathan K., et Anna Di Rienzo. 2010. « Adaptation – Not by Sweeps Alone ». Nature Reviews. Genetics 11 (10): 665 67. https://doi.org/10.1038/nrg2880.
Rietveld, Cornelius A., Sarah E. Medland, Jaime Derringer, Jian Yang, Tõnu Esko, Nicolas W. Martin, Harm-Jan Westra, et al. 2013. « GWAS of 126,559 Individuals Identifies Genetic Variants Associated with Educational Attainment ». Science (New York, N.Y.) 340 (6139): 1467 71. https://doi.org/10.1126/science.1235488.
Rindermann, Heiner, David Becker, et Thomas R. Coyle. 2020. « Survey of Expert Opinion on Intelligence: Intelligence Research, Experts’ Background, Controversial Issues, and the Media ». Intelligence 78 (janvier): 101406. https://doi.org/10.1016/j.intell.2019.101406.
Roth, Philip L., Chad H. Van Iddekinge, Philip S. DeOrtentiis, Kaylee J. Hackney, Liwen Zhang, et Maury A. Buster. 2017. « Hispanic and Asian performance on selection procedures: A narrative and meta-analytic review of 12 common predictors ». Journal of Applied Psychology 102 (8): 1178 1202. https://doi.org/10.1037/apl0000195.
Rushton, J. Philippe, et Arthur R. Jensen. 2005. « “Thirty years of research on race differences in cognitive ability”: Correction to Rushton and Jensen (2005) ». Psychology, Public Policy, and Law 11 (3): 406 406. https://doi.org/10.1037/1076-8971.11.3.406.
Scheiber, Caroline. 2016. « Do the Kaufman tests of cognitive ability and academic achievement display construct bias across a representative sample of Black, Hispanic, and Caucasian school-age children in grades 1 through 12? » Psychological Assessment 28 (8): 942 52. https://doi.org/10.1037/pas0000236.
Sear, Rebecca. 2022. ‘National IQ’ datasets do not provide accurate, unbiased or comparable measures of cognitive ability worldwide. https://doi.org/10.31234/osf.io/26vfb.
Shull, George Harrison. 1948. « What Is “Heterosis”? » Genetics 33 (5): 439 46. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1209417/.
« The Fight for Our National Intelligence / by Raymond B. Cattell. » s. d. Wellcome Collection. Consulté le 11 octobre 2023. https://wellcomecollection.org/works/mnu7kph2.
« View on IQ ». 2019. 1 juillet 2019. https://viewoniq.org/.
Ward, Mary, George McMahon, B. St. Pourcain, David Evans, Cornelius Rietveld, Daniel Benjamin, Philipp Koellinger, David Cesarini, George Davey Smith, et Nicholas Timpson. 2014. « Genetic Variation Associated with Differential Educational Attainment in Adults Has Anticipated Associations with School Performance in Children ». PloS one 9 (juillet): e100248. https://doi.org/10.1371/journal.pone.0100248.
Warne, Russell. 2022. « National Mean IQ Estimates: Validity, Data Quality, and Recommendations ». Evolutionary Psychological Science 9 (décembre). https://doi.org/10.1007/s40806-022-00351-y.
Warne, Russell T. 2020a. « Continental Genetic Ancestry Source Correlates with Global Cognitive Ability Score ». Mankind Quarterly 60 (3). https://doi.org/10.46469/mq.2020.60.3.7.
———. 2020b. In the Know: Debunking 35 Myths about Human Intelligence. Cambridge University Press.
———. 2021. « Between-Group Mean Differences in Intelligence in the United States Are >0% Genetically Caused: Five Converging Lines of Evidence ». The American Journal of Psychology 134 (4): 480 501. https://doi.org/10.5406/amerjpsyc.134.4.0479.
Woodley of Menie, Michael A. 2015. « How fragile is our intellect? Estimating losses in general intelligence due to both selection and mutation accumulation ». Personality and Individual Differences 75 (mars): 80 84. https://doi.org/10.1016/j.paid.2014.10.047.
Crédit photo : DR
[cc] Breizh-info.com, 2023, dépêches libres de copie et de diffusion sous réserve de mention et de lien vers la source d’origine










2 réponses à “Etudes sur le QI.”
si les « asiatiques de l’Est » réussissent mieux, n’est ce pas aussi parce qu’ils travaillent plus que les autres?
Merci pour cet article de synthèse fort intéressant et bien documenté. Mais je n’arrive pas à déterminer qui en est l’auteur avec qui je souhaiterais entrer en contact. Merci.